AXE 2 : ÉCOLOGIE FONCTIONNELLE DES NÉO-ÉCOSYSTÈMES FORESTIERS
Le concept de « nouvel écosystème » (novel ecosystem) a émergé dans les années 2000 pour désigner soit des écosystèmes « naturels » fortement dégradés, soit des infrastructures anthropiques abandonnées, en évolution libre[1]. Ces « néo-écosystèmes » sont caractérisés par des assemblages d’espèces inédits, conséquences directes de forçages anthropiques dans un environnement changeant, et par une dynamique « naturelle » durablement sinon définitivement orientée par les perturbations anthropiques du passé. Dans ce projet, nous nous concentrerons sur les néo-écosystèmes forestiers, et prioritairement, ceux issus d’une invasion biologique, c’est-à-dire dont l’espèce végétale structurante est exotique. D’autre part, nous souhaiterions étudier les modalités de mise en place de néo-écosystèmes forestiers après une destruction complète des écosystèmes forestiers pré-existants. En cohérence avec la thématique et les compétences de l’unité, nous envisageons d’éclaircir les règles d’assemblage et de fonctionnement des néo-écosystèmes forestiers, et ainsi d’identifier les services écosystémiques qui se trouvent altérés et ceux qui apparaissent de novo, en ciblant prioritairement les interactions émergentes avec les systèmes de production (Figure 2).
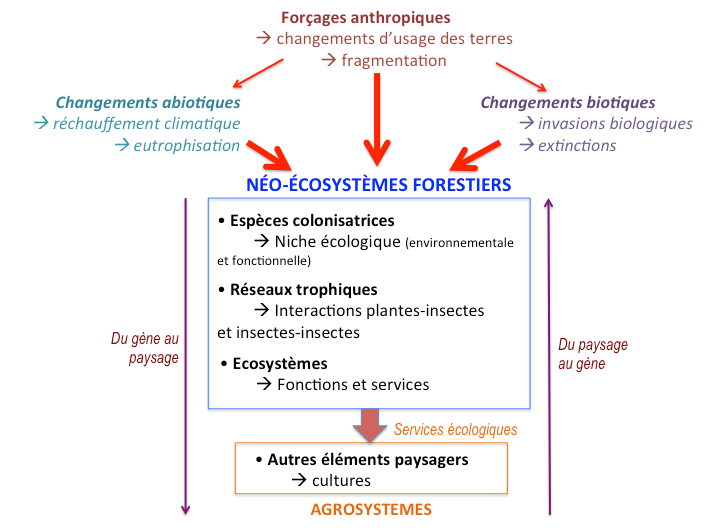
Figure 2 : Schéma de synthèse de l’axe « Ecologie fonctionnelle des néo-écosystèmes forestiers ». En contexte de changements globaux, abiotiques et biotiques, et sous les contraintes propres aux espaces fortement anthropisés comme les systèmes de production, les bois et forêts forment désormais des néo-écosystèmes forestiers, au fonctionnement particulier, délivrant de nouveaux services aux sociétés, notamment en termes de services (positifs ou négatifs) rendus à l’agriculture. Dans le présent projet scientifique, nous cherchons à mieux comprendre le fonctionnement de ces néo-écosystèmes et comment ils interagissent avec les systèmes de production agricoles et forestiers.
-
ASSEMBLAGE ET FONCTIONNEMENT DES NÉO-ÉCOSYSTÈMES
À la fois conséquence et cause de changements locaux, régionaux ou globaux, les invasions biologiques sont devenues une préoccupation majeure à l’échelle mondiale du fait de leurs impacts écologiques, économiques et sociaux. L’écologie des invasions a connu un succès grandissant ces dernières décennies, avec des avancées majeures pour la science, mais de nombreuses inconnues demeurent, notamment sur les conséquences à long terme de certaines plantes invasives qui incorporent durablement des écosystèmes. Les recherches menées ont pour objectif de mieux documenter ces impacts à long terme dans le cas d’écosystèmes forestiers, mais aussi tenter une nouvelle approche pour mieux les anticiper.
Jusqu’alors, la recherche sur les plantes invasives s’est essentiellement concentrée sur les traits d’une espèce qui lui confère sa capacité invasive (invasiveness) et les attributs d’un écosystème qui le rendent vulnérable aux invasions (invasibility). Relativement peu d’études ont porté sur les impacts et encore moins sur les impacts sur le fonctionnement des écosystèmes, c’est-à-dire sur les processus à l’origine des flux d’énergie et des cycles du carbone, de l’azote, de l’eau et des nutriments au sein des écosystèmes. Pourtant, du fait de leur abondance et des nouvelles fonctions écologiques qu’elles apportent, les plantes invasives sont susceptibles de devenir les nouveaux ingénieurs de l’écosystème envahi[2], imposant de nouveaux filtres environnementaux à l’ensemble de l’écosystème et, de manière ultime, créant un « nouvel » écosystème. Nous souhaitons donc explorer les processus qui permettent aux plantes invasives d’interagir avec les espèces résidentes pour former de nouveaux écosystèmes, au fonctionnement original, susceptible de délivrer de nouveaux services aux sociétés. Jusqu’ici, les études sur les plantes invasives se sont concentrées sur les milieux ouverts et des espèces pionnières capables d’envahir des milieux perturbés ; a contrario, les milieux fermés et peu perturbés, comme les forêts, ont été très peu étudiés et même souvent considérés comme résistants aux invasions. Pourtant, il a été montré récemment que si les forêts étaient effectivement touchées par un plus petit nombre d’espèces invasives, le taux de persistance à long terme de ces espèces est beaucoup plus élevé du fait de leur forte tolérance à l’ombre[3]. Etonnamment, les plantes invasives tolérantes à l’ombre n’ont quasiment pas été étudiées alors qu’elles sont susceptibles de déployer des effets à long terme sur le fonctionnement des écosystèmes forestiers. Dans ce projet nous privilégierons donc des espèces ligneuses tolérantes à l’ombre (au moins dans les stades juvéniles), comme le cerisier tardif (Prunus serotina Ehrh.) qui est une espèce modèle bien maîtrisée au sein de l’unité et l’érable sycomore (Acer pseudoplatanus L. indigène en Europe, invasif aux Etats-Unis et en Nouvelle-Zélande). Les travaux de l’unité EDYSAN portent plus particulièrement sur :
– le concept de « pré-adaptation[4] », de « niche vacante » et l’hypothèse du déséquilibre évolutif (Evolutionary Imbalance Hypothesis ou EIH)[5].
– le rôle des interactions gènes-environnement dans le succès invasif.
– une meilleure compréhension des impacts des espèces invasives.
-
INTERACTIONS ÉMERGENTES AVEC LES SYSTÈMES DE PRODUCTION
La drosophile invasive Drosophila suzukii est un insecte asiatique ravageur qui touche les systèmes de production fruitière du nord de la France depuis 2012 et a continué à s’étendre vers l’Europe du nord et de l’est depuis[6]. Sa grande polyphagie et son ovipositeur en forme de scie lui confèrent une niche alimentaire extrêmement large par rapport à ses congénères indigènes ; surtout elle est capable de se nourrir et de pondre sur des fruits immatures, contrairement à ses congénères « indigènes » comme D. melanogaster, d’où son statut de ravageur. Si cet insecte a fait l’objet de nombreuses recherches dans le cadre des productions fruitières, la mise en contexte paysager et l’étude des interactions entre écosystèmes « sauvages » et agrosystèmes, reste assez peu documentée, même dans son aire géographique d’origine[7]. Dans son aire d’origine, D. suzukii est connue pour utiliser des espèces indigènes de cerisier comme hôte : Prunus nipponica et P. donarium. D’autre part, elle est également invasive dans l’aire d’indigénat de P. serotina, aux Etats-Unis. Dans ce contexte, les travaux de l’unité EDYSAN portent sur :
– la niche trophique de D. suzukii et sa dimension spatio-temporelle (i.e., phénologie des fruits sauvages et cultivés) en contexte de réchauffement climatique.
– l’élucidation de la relative résistance de D. suzukii à ses ennemis naturels potentiels, notamment les espèces d’hyménoptères endoparasitoïdes.
[1] Hobbs et al. Global Ecology and Biogeography 2006; 15 ; 1-7. Murcia C et al. Trends in Ecology and Evolution 2014; 29 : 548-543. Hobbs et al. Trends in Ecology and Evolution 2014; 29 : 645-646.
[2] Cuddington K & Hastings A. Ecological Modelling 2004; 178 : 335–347.
[3] Martin PH et al. Frontiers in Ecology and the Environment 2008; 7 : 142-149.
[4] Fridley JD. Annals of the New York Academy of Sciences 2013; 1293, 8–17.
[5] Fridley JD & Sax DF. Global Ecology and Biogeography 2014; 23 : 1157–1166. Vermeij GJ. Science 1991; 253 : 1099–1104.
[6] Cini A et al. Bulletin of Insectology 2012; 65: 149–160. Asplen MK et al. Journal of Pest Science 2015; 88: 469-494.
[7] Mitsui H et al. Entomological Science 2010; 13 : 60–67. Walsh DB et al. Journal of Integrated Pest Management 2011; 1: 1-7. Lee JC et al. Annals of the Entomological Society of America 2015 108: 117-129. Kenis M et al. Journal of Pest Science 2016; doi 10.1007/s10340-016-0755-6. Diepenbrock LM et al. Journal of Pest Science 2016, doi 10.1007/s10340-016-0764-5.